Ongoing major research projects in the lab
Disease and demography of Indigenous Australians before and after European invasion (Nitschke, Bradshaw)

Within the Australian Research Council Centre of Excellence for Indigenous and Environmental Histories and Futures (CIEHF), we are developing models to determine the impacts of diseases derived from European colonists on the demography and cultural landscape of Indigenous Australians (main collaborators: Bird, Griffiths, Ingrey, McNiven, Russell, Saltré, Tobler, Ulm, Williams, Gujaga Foundation).
Future child health under environmental and demographic change (Fatima, Bradshaw)
As a member of the Future Child Health research group, we are modelling the impacts of environmental degradation and rising human densities on the health of future children, both in Australia and globally (main collaborators: Judge, Le Souëf).
Ecological network modelling (Llewelyn, Doherty, Bradshaw)

Ecological network modelling is a method used to understand the interdependence of groups of organisms in an ecosystem. It involves modelling the flow of a biologically relevant currency, such as energy or a particular nutrient, through a system. Importantly, it can be used to model the effects of environmental change on an ecosystem. For example, ecological networks can be used to investigate the cascading effects of the addition of a new species (e.g., invasive species), the loss of a species, or changes in forcing functions (i.e., external factors) such as temperature and rainfall. Network modelling has been widely applied to marine systems, where it is used in resource management to guide the harvesting of fish and other marine organisms. This method can also be used to model ancient extinction events. However, network modelling has rarely been applied in this way. This research aims to use ecological network modelling to understand the late Pleistocene megafauna extinctions. This novel research will provide insight into the processes that could cause extinction patterns similar to those observed in the Late Pleistocene, helping us understand the drivers of megafauna extinction, and the processes responsible for determining the composition of Australia’s current biodiversity (main collaborators: Clement, Cloutier, Long, Saltré, Strona).
Koala demography, genetics, distribution, and genetic rescue in South Australia (Beaman, Bradshaw)

We are predicting population size and trends, genetic structure and constraints, and environmental impacts of the South Australia mainland population of koalas to determine their fate in response to future climate and potential management interventions (main collaborators: Beheregaray, Belov, Burke Da Silva, Hogg, Rogers, Saltré)
Reduction of invasive deer from South Australia (Hamnett, Bradshaw)

We are developing models to estimate the most efficient ways to reduce the densities of feral deer species in South Australia to diminish their ecological and economic impacts (main collaborators: Fielder, Korcz, Page, Saltré)
Estimating the monetary costs of invasive species to human health (Bradshaw)
The InvaHealth programme is supported by the Centre de Synthèse et d’Analyse sur la Biodiversité (Fondation pour la Recherche sur la Biodiversité). It is based on a framework including an eco-epidemiological component detailing various mechanisms affecting health and an economic component assessing cost distribution among stakeholders. This resource will facilitate future research, inform science-based policies, and contribute to better human health outcomes by preventing and addressing invasion-driven impacts, while promoting biodiversity conservation and ecological integrity (main collaborators: Angulo, Courchamp, Courtois, Cuthbert, Diagne, Kourantidou, Latombe, Pontifes, Renault, Roiz, Turbelin).
Modelling the ecology of frogs to guide conservation (Mathwin, Bradshaw)

Together with environmental data, we are examining which conditions signal the reproductive readiness in male and female frogs. We develop population viability models to inform the duration of environmental watering to maximise frog reproductive success in the Murray-Darling Basin (main collaborators: Saltré, Wassens, Ye, Young).
Shark-bite risk and mitigation measures (Riley, Bradshaw)

Maddie Riley is leading this research programme for her PhD thesis. She is testing and development shark-bite mitigation measures and modelling the probability of shark bites under different management interventions (main collaborators: Huveneers).
Palaeo-vegetation models for Australia (Bradshaw)
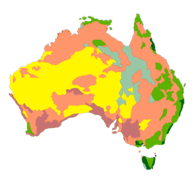
Tracing the role of environmental fluctuations on the evolution of Australia’s natural history has long been hampered by a lack of reasonable reconstructions of vegetation communities over spatial times scales of sufficient magnitude to test hypotheses. For example, to what extent did changing vegetation arising from climate change affect animal extinctions over the Late Pleistocene and early Holocene? How did vegetation communities influence the distribution and movements of ancient human societies? To what extent did climate, fire and grazing interact to alter these same vegetation communities, and what were the implications of these feedbacks on the taxa exploiting them for food and shelter? We are examining these questions through dynamic vegetation models (main collaborators: Saltré).
—–
, it depicts biodiversity scenery from around the world. Visit the Lab to see this 20-m wonder in person.")
